
Статья опубликована в рамках: V Международной научно-практической конференции «Научные достижения биологии, химии, физики» (Россия, г. Новосибирск, 06 марта 2012 г.)
Наука: Биология
Секция: Биохимия
Скачать книгу(-и): Сборник статей конференции
- Условия публикаций
- Все статьи конференции
дипломов
Активность Na+, K+-АТФазы плазматических мембран клеток печени и мозга крыс при хронической алкогольной интоксикации и при введении уксуснокислого цинка
Харченко Ольга Ивановна
канд. биол. наук, мл. науч. сотр., Киевский национальный университет им. Т. Шевченко, г. Киев
E-mail:
Богун Лариса Ивановна
канд. биол. наук, науч. сотр., Киевский национальный университет им. Т. Шевченко, г. Киев
Остапченко Людмила Ивановна
д-р биол. наук, профессор, Киевский национальный университет им. Т. Шевченко, г. Киев
Введение. В основе развития алкоголизма лежат глубокие изменения структуры и функций плазматических мембран различных клеток, которые приводят к нарушению функционирования мембраносвязанных ферментов. Эти энзимы могут выступать в роли биохимических мишеней, которые обуславливают, с одной стороны, повреждающий эффект этанола, а с другой – приспособление организма к алкоголизации.
На сегодня известно, что развитие хронической алкогольной интоксикации сопровождается дефицитом цинка в ряде органов человека и животных. Для коррекции дефицита цинка используют его соли, среди которых низкой токсичностью характеризуется уксуснокислый цинк [8]. Поэтому целью нашей работы было определить влияние уксуснокислого цинка на активность Na+,K+-АТФазы в плазматических мембранах клеток печени и мозга крыс в условиях хронической алкогольной интоксикации.
Материалы и методы. Исследования проводили на самцах белых беспородных крыс массой 180—200 г, которые содержались на стандартном рационе вивария со свободным доступом к воде. 40° этиловый спирт вводился перорально из расчета 2 мл на 100 г массы животного раз в сутки. Животные были разделены на 3 группы. 1-вая группа – контрольные животные; 2-рая – крысы с хронической алкогольной интоксикацией, которая вызывалась по методике М. Х. Халилова и Ш. А. Закихорджаева [7]; 3-тья – крысы с хронической алкогольной интоксикацией, которым дополнительно вводили уксуснокислый цинк в дозе 2 мг на 100 г массы животного [8]. Животных декапитировали на 4, 7, 11, 16 и 21 сутки. Общую фракцию гепатоцитов крыс получали по модифицированной методике [5]. Получение фракций плазматических мембран клеток печени и мозга крыс, а также определение активности Na+,К+-АТФазы проводили согласно [6]. Определение концентрации белка проводили по методу [9]. Статистическую обработку полученных данных проводили общепринятыми методами вариационной статистики на основании 7—12 повторов (M±m, n=7–12) [1].
Результаты. При исследовании действия этанола на активность Na+, K+-AТФазы плазматических мембран гепатоцитов крыс нами было установлено ее возрастание на 4-тые (на 25 %) и 7-мые сутки (на 50 %), снижение на 11-тые сутки (на 36,4 %) и повторное повышение активности фермента на 16-тые (в 2,5 раз) и 21-вые (в 5 раз) сутки эксперимента по сравнению с контролем (рис. 1).
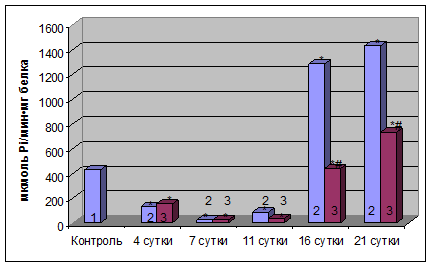
Рис. 1. Активность Na+,К+–АТФазыплазматических мембран гепатоцитов крыс при хронической алкогольной интоксикации и при введении уксуснокислого цинка (1 – контроль; 2 – этанол; 3 – этанол + цинк).
* – Р≤0,05 по сравнению с контролем
#– Р≤0,05 по сравнению с экспериментальной моделью
Введение уксуснокислого цинка при хронической алкогольной интоксикации приводило к возрастанию активности данного фермента плазматических мембран гепатоцитов крыс на 7-мые (на 40 %) и 16-тые сутки (на 15 %) по сравнению с контрольными значениями, в то время как на 4-тые и 11-тые сутки этот показатель не отличался от контроля. Во время последнего этапа исследования (21-вые сутки) активность Na+,К+-АТФазы была в 2,5 раза выше по сравнению с контролем (рис. 1).
Таким образом, уксуснокислый цинк приводит к уменьшению влияния этанола на Na+,К+-АТФазу плазматических мембран гепатоцитов крыс, снижая активность этого фермента. При введении уксуснокислого цинка наблюдается снижение активности Na+,К+-АТФазы по сравнению с соответствующими этапами в условиях алкогольной интоксикации на начальных этапах (на 4-тые и 7-мые сутки – на 20 % и 7 %, соответственно), а также на более поздних сроках эксперимента (на 16-тые и 21-тые сутки – на 54 % и 50 %, соответственно), в то время как на 11-тые сутки эксперимента наблюдается незначительное повышение активности энзима.0
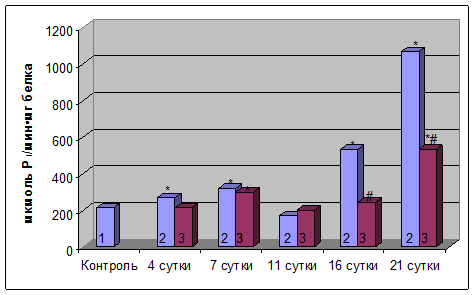
Рис. 2. Активность Na+,К+–АТФазыплазматических мембран клеток мозга крыс при хронической алкогольной интоксикации и при введении уксуснокислого цинка (1 – контроль; 2 – этанол; 3 – этанол + цинк).
* – Р≤0,05 по сравнению с контролем
#– Р≤0,05 по сравнению с экспериментальной моделью
В отличие от гепатоцитов, в клетках мозга при хронической алкоголизации было выявлено снижение активности Na+,K+-AТФазы плазматических мембран на более ранних этапах исследования на 4-тые, 7-мые и 11-тые сутки (в 3, 16 и 5 раз, соответственно) в сравнении с контролем. В дальнейшем наблюдался резкий рост активности фермента на 16-тые и 21-вые сутки эксперимента (в 3 и 3,3 раза, соответственно) по сравнению с контролем (рис. 2).
При хронической алкогольной интоксикации введение уксуснокислого цинка приводило к снижению активности исследуемого фермента плазматических мембран клеток мозга крыс на 4-тые, 7-мые и 11-тые сутки эксперимента по сравнению с контролем (в 3, 20 и 15 раз, соответственно). На 21-вые сутки активность Na+,K+-AТФазы возрастала в 1,8 раз по отношению к контрольным значениям. В сравнении с соответствующими сроками при действии этанола активность энзима на 11-тые, 16-тые и 21-вые сутки была соответственно в 3, 2,8 и 1,8 раз более низкой.
Na+,K+-AТФаза (АТФ-фосфогидролаза, КФ 3.6.1.37) –интегральный белок плазматических мембран клеток, который осуществляет энергозависимое противоположно направленное перенесение ионов Na+ и K+. Na+, K+-AТФаза вовлечена в многочисленные клеточные функции и процессы, связанные с существованием ионных градиентов, в частности в обеспечение электрической возбудимости нервной и мышечной тканей [12]. В то же время показано, что при разных патологических состояниях наблюдается ее инактивация [10]. Причиной этого может быть или прямое действие на фермент, или структурные изменения в мембране (например, в результате свободнорадикальных процессов при церебральной ишемии, сублетальном ионизирующем облучении), или многоуровневые нарушения тканеспецифичных клеточных механизмов регуляции активности и экспрессии изоферментов Na+,K+-AТФазы при разных хронических патологических состояниях [3].
Известно, что этанол вызывает снижение текучести мембран. Согласно данных ряда авторов это сопровождается усилением активного трансмембранного транспорта Na+ в результате увеличения числа переносчиков и роста их родства к этому иону, а также стабилизации внутри- и внеклеточного обмена Ca2+.
Существующие на сегодня данные относительно влияния этанола на активность Na+, K+-AТФазы носят противоречивый характер. Так при изучении прямого действия этанола на Na+, K+-AТФазу в опытах in vitro показано угнетение активности этого фермента [11]. Чувствительность Na+, K+-АТФазы к действию этанола in vitro зависит от целостности белок-липидного комплекса фермента в мембране [4]. Кроме того и от ее фосфолипидного окружения, в частности кислых фосфолипидов, которые обеспечивают формирование негативного заряда на поверхности мембран, чем обусловливают отталкивание между последними и притягивание поликатионних белков [2]. Полученные нами данные о содержании фосфолипидов при влиянии этанола и их коррекция при введении уксуснокислого цинка может быть одним из факторов показанных нами изменений активности фермента.
Выводы. Таким образом, нами показано, возрастание активности Na+, K+-AТФазы плазматических мембран клеток печени и мозга крыс в динамике развития хронической алкогольной интоксикации. Введение уксуснокислого цинка приводило к снижению активности Na+, K+-AТФазы. На основании полученных нами результатов, можно сделать вывод о перспективности его дальнейшего изучения с целью использования для коррекции метаболических нарушений, что может быть основой разработки новых лечебных противоалкогольных средств.
Список литературы:
- Брандт З. Статистические методы анализа наблюдений. // М.: Мир, 1975. – 312 с.
- Горчев В. Ф. Роль биологических мембран в энтропийных процессах живых организмов // Інформаційна та негентропійна терапія. – 1995. –№ 2. –С. 4—9.
- Капля А. А. Структурная организация и функциональная роль изоферментов Na+,K+-АTPазы // Киев: Киевский университет, 1998. – 162 с.
- Мищук Д. О., Капля А. А. Влияние этанола на структурно-функцинальные характеристики мембран коры головного мозга крыс in vitro // Укр. біохім. журн. – 2003. – Т. 75, № 2. – С. 55—61.
- Петренко А. Ю., Сукач А. Н., Росляков А. Д. Выделение гепатоцитов крыс неферментативным методом: детоксикационная и дыхательная активность // Биохимия. – 1991. - Т. 56. - Вып. 9. – С. 1647—1650.
- Рыбальченко В. К.,Коганов М. М. Структура и функции мембран. Практикум – К.: „Вища школа” – 1988. – 254 c.
- Халилов М. Х., Закихорджаев Ш. Я. К характеристике некоторых патохимических сдвигов в крови, тканях печени и головного мозга при экспериментальной алкогольной интоксикации // Вопросы клиники алкоголизма: Сб. науч. тр., Ташкент, 1983. – С. 38—41.
- Ещенко Ю. А. Вміст цинку в клітинах при різних функціональних станах інсулярного апарата підшлункової залози: автореф. дис. на здобуття наук. ступеня канд. біол. наук: спец. 03.00.13 «Фізіологія людини і тварин» / Ю. А. Ещенко. – Київ, 2004. – 19 с.
- Bradford M. M.A rаpid and sensitive method for quantities of utilizing the principle of protein binding // Analytical Biochemistry. – 1976. – V. 86. – P. 193—200.
- Jamme I., Barbey O., Trouvé P. et al. Focal cerebral ischaemia induces a decrease in activity and a shift in ouabain affinity of Na+,K+-ATPase isoforms without modifications in mRNA and protein expression // Brain Res.– 1999. –V. 20, № 819(1—2). – P. 132—142.
-
11.Omodeo-Salé F., Lindi C., Palestini P., Masserini M. Role of phosphatidylethanol in membranes. Effects on membrane fluidity, tolerance to ethanol, and activity of membrane-bound enzymes // Biochemistry.– 1991. –V. 5, № 30(9). – P. 2477—2482.
-
Scheiner-Bobis G. The sodium pump. Its molecular properties and mechanisms of ion transport // Eur J Biochem. – 2002. – V. 269. – P. 2424—2433.
дипломов
Оставить комментарий