
Статья опубликована в рамках: X Международной научно-практической конференции «Научное сообщество студентов XXI столетия. ЕСТЕСТВЕННЫЕ НАУКИ» (Россия, г. Новосибирск, 06 мая 2013 г.)
Наука: Биология
Секция: Экология
Скачать книгу(-и): Сборник статей конференции
- Условия публикаций
- Все статьи конференции
дипломов

ПОЛИГОНАЛЬНАЯ ВИЗУАЛИЗАЦИЯ МНОГОМЕРНЫХ ДАННЫХ И РЕГРЕССИОННЫЙ АНАЛИЗ В МЕТОДОЛОГИИ ИССЛЕДОВАНИЙ ХИРОНОМИД НА ПРИМЕРЕ GLYPTOTENDIPES GLAUCUS MG
Шмаров Вячеслав Анатольевич
магистрант 2 года обучения направления «Биология» по программе «Экология», кафедра биоэкологии и биоразнообразия химико-биологического института БФУ им. И. Канта, г. Калининград
E-mail: enant@list.ru
Винокурова Наталья Владимировна
научный руководитель, канд. биол. наук, доцент кафедры биоэкологии и биоразнообразия БФУ им. И. Канта, г. Калининград
Актуальность темы
В связи с увеличением темпов роста хозяйственной деятельности человека неминуемо возрастают и масштабы негативного антропогенного воздействия на окружающую среду, последствия которых стохастичны и поэтому крайне опасны. В настоящее время к приоритетным ксенобиотикам относятся тяжелые металлы, играющие значительную роль и в загрязнении водоемов.
Общеизвестно, что личинки семейства Chironomidae являются доминирующими видами макрозообентоса. Хирономиды имеют самые крупные в природе политенные хромосомы, которые позволяют проводить цитогенетический анализ воздействия различных токсических веществ на хромосомные перестройки, вследствие чего хирономиды представляют собой удобную модель при анализе антропогенных воздействий. Также хирономиды способны легче, чем другие водные организмы, накапливать тяжелые металлы внутри своего тела благодаря высокой проницаемости покровов [8].
Многие исследователи для биомониторинга водоёмов предлагают использовать в качестве биоиндикаторов личинок хирономид [1, 2, 5, 10, 12—16]. Большую часть аспектов использования хирономид на организменном, популяционном, экосистемном уровне биомониторинга детально рассматривает Т.Д. Зинченко [7]. Однако вопрос о воздействии тяжелых металлов на бентосные организмы и, в частности, на личинок хирономид на сегодняшний день изучен недостаточно. Одним из препятствий является определение специфичности воздействия отдельного элемента в целом комплексе токсических веществ, встречающихся в водоёмах, тем более, что показатели могут разниться в зависимости от сезона генерации и половой принадлежности особей и др. [9].
На сегодняшний день перед экологической наукой стоит задача описания принципов, упрощений и абстракций, к которым необходимо научиться сводить многообразие реального мира природы, а также необходимость построения математических моделей, отражающих сложную систему взаимосвязей в природе. В этом случае использование инструментов математического аппарата, методы графического представления многомерных данных и их моделирования можно применять в качестве специфических методов системной экологии, с помощью которых исследуются законы функционирования и развития экосистем во времени и пространстве [6].
Цель работы: выявить характер зависимости изменчивости хромосомного полиморфизма природных популяций Glyptotendipes glaucus Mg. нескольких водоёмов г. Калининграда от сезона генерации и половой принадлежности, а также экспериментальных выборок данного вида хирономид от содержания тяжелых металлов в экспериментальных растворах.
Материалы и методы
Материалом исследования послужили первичные данные цитогенетического анализа политенных хромосом личинок хирономид G. glaucus IV стадии развития, собранных из 6-ти водоёмов г. Калининграда (табл. 1), а также — личинок хирономид G. glaucus IV стадии развития, выращенных в эксперименте из кладок, собранных в прибрежной зоне тех же водоемов с погруженных в воду частей растений, разлагающихся макрофитов и коряг. Даты, места сборов и количество исследованных особей представлены в таблице 1.
Таблица 1.
Места сбора личинок и количество исследованных особей
|
Название водоёма |
Год сбора |
Генерация |
Количество особей |
Количество самцов и самок |
|
Система прудов Карасевка |
2006 |
летняя |
42 |
♀-11, ♂-16, н.о.-15 |
|
2007 |
зимняя |
12 |
♀-5, ♂-2, н.о.-5 |
|
|
2007 |
летняя |
24 |
♀-17, ♂-4, н.о.-3 |
|
|
2008 |
зимняя |
15 |
♀-7, ♂-6, н.о.-2 |
|
|
2008 |
летняя |
45 |
♀-29, ♂-15, н.о.-1 |
|
|
Озеро Пеньковое |
2007 |
летняя |
53 |
♀-31, ♂-12, н.о.-10 |
|
2008 |
зимняя |
43 |
♀-11, ♂-12, н.о.-20 |
|
|
2009 |
летняя |
46 |
♀-18, ♂-17, н.о.-11 |
|
|
Пруд Чистый |
2007 |
летняя |
46 |
♀-26, ♂-15, н.о.-5 |
|
2008 |
зимняя |
10 |
♀-7, ♂-2, н.о.-1 |
|
|
2008 |
летняя |
50 |
♀-37, ♂-4, н.о.-9 |
|
|
Пруд Ботанического сада РГУ |
2008 |
зимняя |
30 |
♀-18, ♂-10, н.о.-2 |
|
Озеро Школьное |
2008 |
зимняя |
22 |
♀-6, ♂-9, н.о.-7 |
|
2008 |
летняя |
14 |
♀-9, ♂-4, н.о.-1 |
|
|
Пруд Нижний |
2009 |
летняя |
51 |
♀-21, ♂-27, н.о.-3 |
Для изучения воздействия металлов на политенные хромосомы G. glaucus в лабораторных условиях были выбраны вещества — сульфат цинка (II), сульфат меди (II), сульфат железа (II), в концентрациях равных концентрациям металлов обнаруженных в грунтах исследуемых природных водоемах города Калининграда и пересчитаны на молярную концентрацию металла.
Собранный материал погружали на месте в растворы FeSO4, ZnSO4 и CuSO4, а также в отстоянную воду (контроль) (2—3 кладки на концентрацию). После выклева (2—3-й день) личинок перемещали в отстоянную воду (личинки из 1 кладки на 1,5 литра воды). Кормление осуществлялось 1 раз в сутки гидролизатом дрожжей. Личинки выращивались до 4-го возраста в течение 2—4 месяцев, после чего фиксировались в спиртоуксусной смеси [3, 4].
Математическая обработка и визуализация данных проводилась с помощью методов профилограмм и регрессионного анализа с использованием пакета программ STATISTICA 5.0.
Всего из природных популяций Glyptotendipes glaucus было исследовано 503 особи. Из экспериментального материала было исследовано 60 особей G. glaucus в контроле, и 380 особей из различных концентраций исследуемых веществ.
Результаты и обсуждение
Число исследованных особей, число инверсионных последовательностей и параметры хромосомного инверсионного полиморфизма личинок хирономид в анализируемых водоемах представлены в табл. 2.
Таблица 2.
Общие показатели инверсионного полиморфизма исследованных популяций G.glaucus города Калининграда
|
Пруд Ботсада |
Озеро Школьное |
Озеро Пеньковое |
Пруд Нижний |
пруды Карасевка |
Пруд Чистый |
|
|
Число исследованных особей |
30 |
36 |
142 |
51 |
138 |
106 |
|
Число инверсионных последовательностей |
13 |
13 |
21 |
16 |
19 |
17 |
|
Число геномных комбинаций |
22 |
16 |
32 |
18 |
26 |
19 |
|
Отношение числа инверсионных последовательностей к числу геномных комбинаций (ЧИП/ЧГК) |
0,59 |
0,84 |
0,86 |
0,88 |
1,14 |
1,6 |
Анализ материала по особям мужского и женского пола дает следующие результаты (табл.3).
Таблица 3.
Основные показатели инверсионного полиморфизма самцов и самок G.glaucus из природных популяций водоемов г. Калининграда
|
Показатели полиморфизма |
Оз. Карасевка n=112 |
Оз. Пенько-вое n=101 |
Пруд Чистый n=91 |
Оз. Школьное n=28 |
Пруд Нижний n=48 |
Пруд Ботани-ческого сада n=28 |
||||||
|
♂ |
♀ |
♂ |
♀ |
♂ |
♀ |
♂ |
♀ |
♂ |
♀ |
♂ |
♀ |
|
|
Число особей |
43 |
69 |
41 |
60 |
21 |
70 |
13 |
15 |
27 |
21 |
10 |
18 |
|
Число инверсионных последовательностей |
14 |
16 |
12 |
17 |
10 |
13 |
11 |
12 |
13 |
12 |
11 |
13 |
|
Число геномных комбинаций |
11 |
20 |
18 |
21 |
4 |
11 |
6 |
9 |
12 |
11 |
9 |
14 |
|
Отношение числа инверсионных последовательностей к числу геномных комбинаций (ЧИП/ЧГК) |
1,27 |
0,8 |
0,66 |
0,80 |
2,5 |
1,18 |
1,83 |
1,33 |
1,08 |
1,09 |
1,2 |
0,92 |
Популяция, находясь в условиях стресса, начинает формировать большее количество геномных комбинаций из имеющихся последовательностей дисков хромосом, а увеличение числа инверсионных последовательностей не так велико, что и приводит к закономерному уменьшению показателя отношения числа инверсионных последовательностей к числу геномных комбинаций (ЧИП/ЧГК).
Интерпретация данных с помощью метода профилограмм позволяет собрать воедино и визуализировать данные о влиянии половой дифференциации (рис. 1) и сезона генерации (рис. 2) особей для каждого из исследуемых озер на одном графике-полигоне.
Отношение числа инверсионных последовательностей к числу геномных комбинаций в среднем ниже у самок Glyptotendipes glaucus по сравнению с самцами (1,02 и 1,42 соответственно), что может быть связано с более высокими генетическими адаптационными способностями особей женского пола (рис. 1).

Рисунок 1. Профилограммы значений отношения числа инверсионных последовательностей к числу генотипических комбинаций (ЧИП/ЧГК) для особей Glyptotendipes glaucus мужского и женского пола системы водоемов г. Калининграда
Показатель отношения числа инверсионных последовательностей к числу геномных комбинаций у особей летней генерации в среднем ниже, чем у особей зимней (1,10 и 1,29 соответственно), что может быть связано с более интенсивным процессом формирования геномных комбинаций летом в виду повышения температуры окружающей среды (рис. 2).
Более высокие летние показатели температуры интенсифицируют биохимические процессы в организме, что, в свою очередь, приводит к известным процессам декомпактизации, деконденсации дисков политенных хромосом, делая их более доступными для мутагенных лигандов окружающей природной среды, которые, могут индуцировать хромосомные перестройки.

Рисунок 2. Профилограммы значений отношения числа инверсионных последовательностей к числу генотипических комбинаций (ЧИП/ЧГК) для зимней и летней генераций особей Glyptotendipes glaucus системы водоемов г. Калининграда
В таблице 4 обобщены и приведены основные параметры и данные эксперимента по определению воздействия ионов тяжелых металлов на показатели инверсионного полиморфизма личинок Glyptotendipes glaucus, графическая интерпретация эксперимента отражена на рисунках 3—5.
Таблица 4.
Параметры хромосомного инверсионного полиморфизма экспериментальных личинок Glyptotendipes glaucus в различных концентрациях сульфатов цинка, меди, железа
|
Показатели полиморфизма |
Кон-троль |
Концентрации Zn2+ (ммоль/л) |
Концентрации Cu2+ (ммоль/л) |
Концентрации Fe2+ (ммоль/л) |
|||||
|
0,52 |
0,62 |
0,94 |
0,00945 |
0, 01889 |
0,82 |
1,40 |
7,76 |
||
|
Количество исследованных особей |
60 |
60 |
40 |
40 |
60 |
40 |
40 |
60 |
40 |
|
Число инверсионных последовательностей |
10 |
9 |
10 |
11 |
10 |
11 |
10 |
10 |
11 |
|
Число геномных комбинаций |
5 |
6 |
7 |
11 |
8 |
11 |
6 |
9 |
16 |
|
Отношение числа инверсионных последовательностей к числу геномных комбинаций |
2,0 |
1,5 |
1,42 |
1,0 |
1,25 |
1,0 |
1,66 |
1,11 |
0,68 |

Рисунок 3. Зависимость отношения числа инверсионных последовательностей к числу геномных сочетаний в экспериментальных выборках G. Glaucus от концентраций цинка в эксперименте

Рисунок 4. Зависимость отношения числа инверсионных последовательностей к числу геномных сочетаний в экспериментальных выборках G. glaucus от концентраций меди в эксперименте
Как видно из рисунков 3—5, показатель числа инверсионных последовательностей в концентрациях железа сохранялся на уровне контроля и не показал значимости, как и в растворах цинка и меди. Однако число геномных комбинаций, сформированных этими последовательностями, резко возрастает в 2—3 раза вместе с увеличением концентраций железа в экспериментальном растворе.
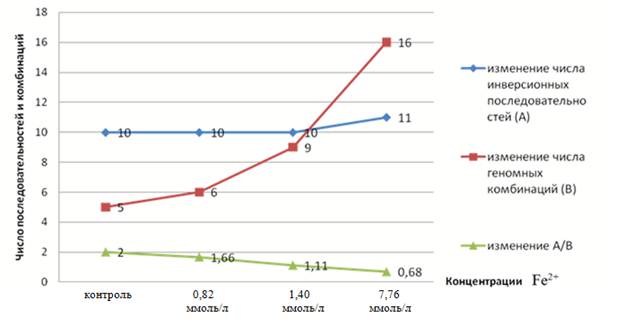
Рисунок 5. Зависимость отношения числа инверсионных последовательностей к числу геномных сочетаний в экспериментальных выборках G. glaucus от концентраций железа в эксперименте
То есть этот показатель изменялся в соответствии с подобной тенденцией в средах с цинком и медью. Также прослеживается уменьшение отношения числа инверсионных последовательностей к числу геномных комбинаций от 1,2 до 3,0 раз вместе с возрастанием концентраций вещества. Подобная закономерность так же наблюдалась в растворах цинка и меди.
Видно, что параметр оценки хромосомного полиморфизма — отношение числа инверсионных последовательностей к числу геномных комбинаций уменьшается в соответствие с увеличением концентраций тяжелых металлов в экспериментальных растворах. Самое высокое значение этого показателя (2,0) наблюдалось в контрольных выборках, а самые низкие показатели — в высоких концентрациях исследуемых металлов.
Регрессионный анализ данных зависимости ЧИП/ЧГК от концентрации металла позволяет получить аналитическое выражение для данной функции, а также оценить скорость уменьшения ЧИП/ЧГК под действием различных металлов и сравнить эти показатели в ряду металлов Zn2+, Сu2+, Fе2+ . (табл. 5, рис. 6)
Таблица 5.
Параметры регрессионного анализа данных зависимости ЧИП/ЧГК от концентрации цинка, меди и железа в эксперименте
|
Аргумент |
Аналитическое выражение функции, ЧИП/ЧГК = f([Ме2+])
|
Первая производная функции
|
Скорость уменьшения ЧИП/ЧГК с увеличением концентрации иона металла, м3/моль |
|
[Zn2+], ммоль/л |
-1.2202 x + 2.1527 |
-1.2202 |
1,22 |
|
[Сu2+], ммоль/л |
-26.4811 x + 1.5002 |
-26.4811 |
26,48 |
|
[Fе2+], ммоль/л |
-0.1108 x + 1.5186 |
-0.1108 |
0,11 |

Рисунок 6. Графики функции ЧИП/ЧГК = f([Ме2+]) в эксперименте для цинка, меди и железа
Как известно, первая производная зависимой величины от аргумента есть скорость изменения данной величины с увеличением аргумента. Скорость уменьшения ЧИП/ЧГК от концентрации металла равна производной функции ЧИП/ЧГК от [Ме2+], взятой с противоположным знаком. Таким образом, скорость уменьшения ЧИП/ЧГК с увеличением концентрации ионов металла возрастает в ряду Fе2+ → Zn2+ → Сu2+ (рис. 6), что означает увеличение степени хромосомного инверсионного полиморфизма в этом ряду и может характеризовать нарастание потенциальной мутагенной активности в данном ряду металлов в отношении политенных хромосом Glyptotendipes glaucus Mg.
Итак, использование метода регрессионного анализа для аналитической интерпретации воздействия ряда концентраций сульфатов меди, цинка, железа на хромосомный инверсионный полиморфизм Glyptotendipes glaucus Mg. в эксперименте позволяет определить градацию потенциальной мутагенной активности в ряду исследуемых ионов тяжелых металлов. Из полученных данных заметно, что потенциальная мутагенная активность металлов возрастает в ряду Fе2+ → Zn2+ → Сu2+, причём в этом ряду наблюдается закономерное уменьшение радиуса иона [11]. Можно предположить, что в живые ткани более интенсивно проникают и кумулируются в них катионы металлов с меньшим радиусом, вызывая хромосомные аберрации.
Заключение
Отношение числа инверсионных последовательностей к числу геномных комбинаций является параметром оценки хромосомного полиморфизма; зависимость этих величин друг от друга носит обратно пропорциональный характер.
Принадлежность особи к женскому полу, летней генерации, а также возрастающая концентрация ионов металлов в среде (в ряду Fe, Zn, Cu) вносят вклад в уменьшение отношения числа инверсионных последовательностей к числу геномных комбинаций, увеличивая уровень хромосомного инверсионного полиморфизма Glyptotendipes glaucus Mg.
Визуализация многомерных данных при помощи метода профилограмм, а также регрессионный анализ экспериментальных выборок составляют предмет интеграции методов системной экологии в методологию исследований хирономид и в случаях ограниченного количества данных, зачастую недостаточных для умозрительного заключения, позволяют анализировать и сравнивать показатели и характеры зависимостей, выявляя закономерности процессов.
Список литературы:
1.Балушкина Е.В. Структура сообществ донных животных и оценка экологического состояния р. Ижоры: влияние гидрофизических и гидрохимических параметров воды // Биология внутренних вод. 2003. № 1. С. 74—80.
2.Балушкина Е.В. Функциональное значение личинок хирономид в континентальных водоемах. Л.: Наука, 1987. 179 с.
3.Винокурова Н.В., Данилова М.В., Сидорова В.В. // Влияние оксида ванадия на конъюгацию гомологов политенных хромосом кариотипа клеток слюнных желёз Chironomus plumosus // Материалы международной научно-технической конференции «Наука и образование». — 2004. — ч. 6. — 167 — 170 с.
4.Винокурова Н.В., Данилова М.В., Шартон А.Ю. К методике выращивания хирономид в лабораторных условиях и их использование в токсикологических экспериментах//Теоретические и прикладные аспекты экологии и биологии: Межвуз. сб. ст./Калининград: Изд-во РГУ им. И. Канта,2008. — с. 77—79.
5.Винокурова Н.В., Червоткина Т.А. Новые данные по фауне хирономид (Diptera: Chironomidae) водоёмов Калининградской области // Вестник Балтийского федерального университета им. И. Канта. 2011. Вып. 7. С. 103—112.
6.Дулепов В.И., Лескова О.А., Майоров И.С. Системная экология. — Владивосток: ВГУЭС, 2004. — 252 с.
7.Зинченко Т.Д. Биоиндикация как поиск информативных компонентов водных экосистем (на примере хирономид — Diptera, Chironomidae) // Чтения памяти Владимира Яковлевича Леванидова. 2005 Вып. 3 , С. 338 — 359.
8.Кикнадзе И.А., Шилова А.И., Керкес И.Е. и др. Кариотипы и морфология личинок трибы Chironomini. Атлас. Новосибирск: Наука, 1991. — 115 с.
9.Майстренко В.Н., Хамитов Р.З., Будников Г.К. Эколого-аналитический мониторинг суперэкотоксикантов. — М.: Химия, 1996. — 319 с.
10.Сергеева И.В., Аникина М.А. Личинки хирономид Procladius (Diptera, Chironomidae) как биоиндикаторы загрязнения реки Волги у Саратова и её притоков с большой антропогенной нагрузкой // Вестник Саратовского госагроуниверситета им. Н.И. Вавилова. 2009. № 4. С. 26—28.
11.Рабинович В.А., Хавин З.Я. Краткий химический справочник, 3-е изд., перераб. и доп. — Л.: Химия, 1991. — 432 с.
12.Hirvenoja M., Michailova P. Chironomus melanotus Keyl and its taxonomical and ecological relations to C. riihimakiensis Wülker (Diptera, Chironomidae) // Spixiana. 1991. V. 14 (3) P. 309—320.
13.Johnson R.K., Wiederholm T., Rosenberg D.M. Freshwater biomonitoring using individual organisms, populations and species assemblages of benthic macro-invertebrates // Freshwater biomonitoring and benthic macroinvertebrates / eds Rosenberg D.M., Resh V.H. N.Y.: Chapman and Hall, 1993. P. 40—158.
14.Thorat L., Bimalendu N. Trends in Chironomid Research //Chironomus Newsletter on Chironomidae Research. 2010. № 23. P. 34—35.
15.Warwick W.F. Morphological deformities in larvae of Procladius Scuse (Diptera: Chironomidae) and their biomonitoring potential // Can. J. Fish. Aquat. Sci. 1989. V. 46. P. 1255—1271.
16.Wiederholm T. Chironomids as indicators of water quality in Swedish lakes// Naturvardsverkets Limnologistika Undersokningar. 1976. V. 10. P. 1—17.
дипломов
Оставить комментарий